生物科技有限公司")




- 手机:13761418683
联系人:爱必信
网址:http://www.absin.cn/
地 址:上海市浦东新区新浩路58号18号楼

激动剂抑制剂全推荐——Kinesin 抑制剂
2021-05-14
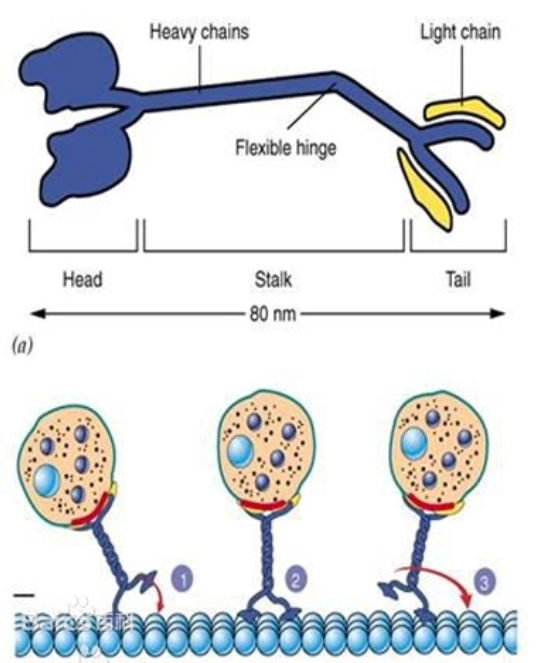
驱动蛋白(Kinesin)是从鱿鱼的轴质(axonplasm)中分离的一种发动机蛋白。由几个不同的结构域组成, 包括两条重链和两条轻链,具有两个球形的头(具有ATP酶活性)、一个螺旋状的杆和两个扇子状的尾。通过结合和水解ATP,导致颈部发生构象改变,使两个头部交替与微管结合,从而沿微管“行走”,将“尾部”结合的“货物”(运输泡或细胞器)转运到其它地方。
KLCs可能通过尾巴增强对Kinesin-1的抑制
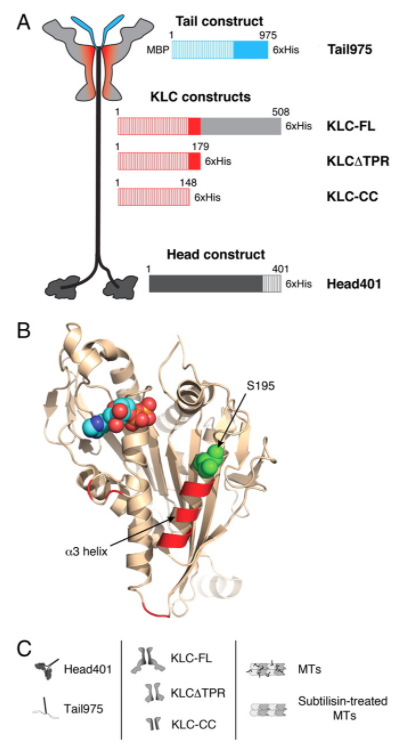
KLCs先前被认为是Kinesin-1活性的调节因子。在细胞提取物和活细胞中,KHC/KLC复合物对MT的结合明显弱于单独的KHC。研究人员提出,KLCs可能通过尾巴增强对Kinesin-1的抑制。研究人员通过使用荧光各向异性直接测量KLCs对头尾结合亲和力的影响。使用突变果蝇KHC头二聚体,没有表面暴露的Cys残基。通过引入S195C突变,用荧光素-5-马来酰亚胺对Head401进行特异性位点标记。S195靠近尾与头的结合位置(14,33),而头尾相互作用导致了容易检测到的各向异性变化。
KLCs抑制Tail-MT结合
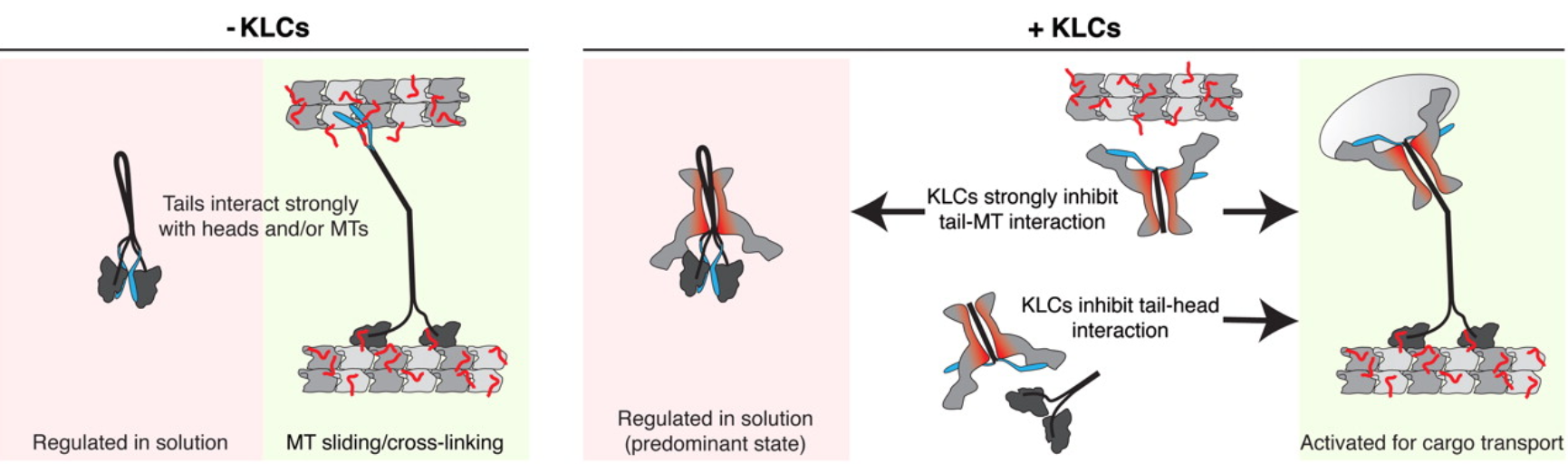
没有KLCs(左),Kinesin-1要么在溶液中被调节,要么在尾部系紧的情况下与MTs结合。由于尾部对头部/MTs的高亲和力,电机不能访问其货物运输胜任状态。在KLCs存在时(右),头尾和尾- mt相互作用被抑制。强烈抑制尾- mt结合意味着受调控的kinesin-1构象成为主要形式,但尾-头亲合力也降低,使马达处于平衡状态,易于被激活用于货物运输。
运动神经元的长轴突运输
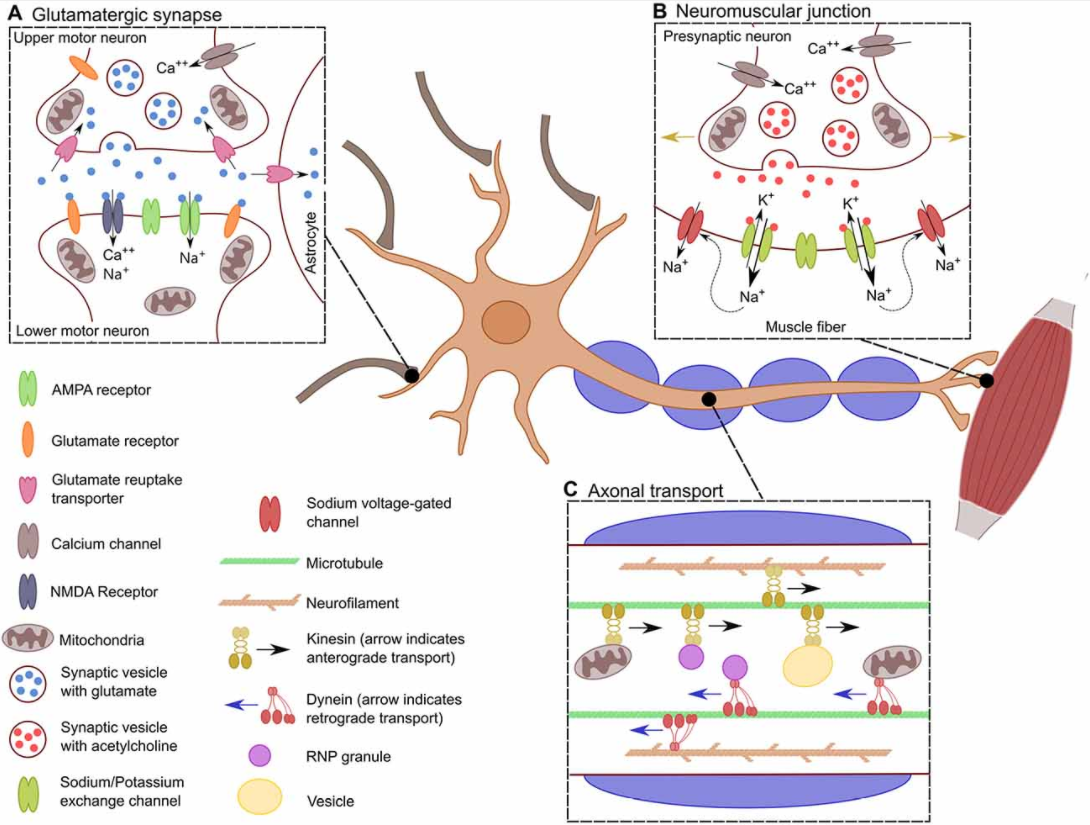
神经元必须根据细胞需要不断地将线粒体、囊泡和RNPs颗粒转移到轴突的局部位置。轴突的转运是双向的,驱动蛋白提供顺行转运,动力蛋白提供逆行转运。这对蛋白质、转录本和细胞器的适当分布是至关重要的。
产品信息:
| 货号 | 名称 | 规格 |
| abs817763 | ARQ 621 | 5mg/25mg |
| abs813933 | ARRY-520 | 5mg/10mg |
| abs811599 | ARRY-520 R enantiomer | 10mg/50mg/5mg |
| abs820804 | AZ82 | 5mg |
| abs810468 | CW069 | 5mg/10mg/50mg/100mg |
| abs810096 | Eg5 Inhibitor III(Dimethylenastron) | 5mg/10mg/50mg/100mg |
| abs812254 | EMD534085 | 5mg/10mg |
| abs810313 | GSK-923295 | 5mg/10mg/50mg/100mg |
| abs814576 | H-Cys(Trt)-OH | 100mg/1g |
| abs810098 | K 858 | 5mg/10mg/50mg/100mg |
| abs818775 | Kif15-IN-1 | 2mg/5mg |
| abs818776 | Kif15-IN-2 | 2mg/5mg |
| abs814178 | Litronesib | 100mg/50mg |
| abs818643 | Litronesib Racemate | 5mg/2mg |
| abs811129 | Monastrol | 5mg/10mg/50mg |
| abs810360 | SB-715992 (Ispinesib) | 5mg/10mg/50mg/100mg |
| abs810556 | SB743921 | 5mg/10mg/50mg/100mg |
|
Absin特色产品线(全部现货):
|
- 手机:13761418683